合成生物学論文メモ (May 2021)
読み流した論文のメモ。黒色はメモ、緑色は感想、赤色は特に面白いと思ったもの。
21報
Synthetic Biology
Transcription/Translation Control
遺伝子回路、転写翻訳制御など- The transcriptional landscape of a rewritten bacterial genome reveals control elements and genome design principles
- Authors: van Kooten, Mariëlle J F M; Scheidegger, Clio A; Christen, Matthias; Christen, Beat
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-23362-y
- Institution: ETH Zürich, Switzerland
ボトムアップな方法でゲノム全体の発現制御様式に示唆を与えていて良い。
- Synthetic neural-like computing in microbial consortia for pattern recognition
- Authors: Li, Ximing; Rizik, Luna; Kravchik, Valeriia; Khoury, Maria; Korin, Netanel; Daniel, Ramez
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-23336-0
- Institution: Israel Institute of Technology, Israel
言いたい事は分かるが無理矢理感が否めない。
- Scalable recombinase-based gene expression cascades
- Authors: Kim, Tackhoon; Weinberg, Benjamin; Wong, Wilson; Lu, Timothy K
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-22978-4
- Institution: MIT, USA
回路設計アイデアが面白い。
- Control of Multigene Expression Stoichiometry in Mammalian Cells Using Synthetic Promoters
- Authors: Patel, Yash D; Brown, Adam J; Zhu, Jie; Rosignoli, Guglielmo; Gibson, Suzanne J; Hatton, Diane; James, David C
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00643
- Institution: University of Sheffield, UK
定性的には興味深い内容だが、定量化部分がやや雑な気も。
RNA Synthetic Biology
アプタマー、リボザイムなど- High-Performance Allosteric Conditional Guide RNAs for Mammalian Cell-Selective Regulation of CRISPR/Cas
- Authors: Hochrein, Lisa M; Li, Heyun; Pierce, Niles A
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.1c00037
- Institution: California Institute of Technology, USA
Toehold switchのデザインをcrRNAに適用したような感じで面白い。
Cell-free / Reconstruction
無細胞系、再構成など- In Vitro Protein Synthesis in Semipermeable Artificial Cells
- Authors: Van Raad, Damian; Huber, Thomas
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.1c00044
- Institution: Australian National University, Australia
発現効率を向上させる余地はまだありそう。
Optogenetics
光駆動型ツール、蛍光イメージング、光受容体など、その他〇〇genetics系- Circularly permuted LOV2 as a modular photoswitch for optogenetic engineering
- Authors: He, Lian; Tan, Peng; Zhu, Lei; Huang, Kai; Nguyen, Nhung T; Wang, Rui; Guo, Lei; Li, Ling; Yang, Yuhan; Huang, Zixian; Huang, Yun; Han, Gang; Wang, Junfeng; Zhou, Yubin
- Journal: Nature Chemical Biology
- Year: 2021
- DOI: 10.1038/s41589-021-00792-9
- Institution: Texas A&M University, USA
LOVドメインの使い辛さを比較的真似しやすい方法で解消していて影響が大きそう。
Protein Engineering
タンパク質工学- Creating RNA Specific C-to-U Editase from APOBEC3A by Separation of Its Activities on DNA and RNA Substrates
- Authors: Tang, Guiyue; Xie, Bingran; Hong, Xiangna; Qin, Hang; Wang, Jingfang; Huang, Hai; Hao, Pei; Li, Xuan
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00627
- Institution: Shanghai University, China
変異を加える残基の選定については書かれていたが、どうやって変異後のアミノ酸を選んだかはよく分からなかった。
CRISPR/Cas
クリスパー系- Optimized CRISPR tools and site-directed transgenesis towards gene drive development in Culex quinquefasciatus mosquitoes
- Authors: Feng, Xuechun; López Del Amo, Víctor; Mameli, Enzo; Lee, Megan; Bishop, Alena L; Perrimon, Norbert; Gantz, Valentino M
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-23239-0
- Institution: UCSD, USA
- CopyCatchers are versatile active genetic elements that detect and quantify inter-homolog somatic gene conversion
- Authors: Li, Zhiqian; Marcel, Nimi; Devkota, Sushil; Auradkar, Ankush; Hedrick, Stephen M; Gantz, Valentino M; Bier, Ethan
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-22927-1
- Institution: UCSD, USA
Metabolic/Signal Pathway Engineering
シグナル経路、代謝経路、酵素工学など- High-Throughput Screening of an Octanoic Acid Producer Strain Library Enables Detection of New Targets for Increasing Titers in Saccharomyces cerevisiae
- Authors: Baumann, Leonie; Bruder, Stefan; Kabisch, Johannes; Boles, Eckhard; Oreb, Mislav
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00600
- Institution: Goethe University Frankfurt, Germany
DNA / Biophysics
DNA、核酸論理回路、ナノスケール構造物など- Patterning Microtubule Network Organization Reshapes Cell-Like Compartments
- Authors: Bermudez, Jessica G; Deiters, Alexander; Good, Matthew C
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00575
- Institution: University of Pennsylvania, USA
自己組織化を誘導するタイプの設計は難しそうだがワクワクする。
- Functional Comparison of Laboratory-Evolved XNA Polymerases for Synthetic Biology
- Authors: Medina, Esau; Yik, Eric J; Herdewijn, Piet; Chaput, John C
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.1c00048
- Institution: Rega Institute for Medical Research, Belgium
複数のツールを比較評価する良い論文。
Alternative Hosts / Strain Engineering
宿主や系統株の開発- Programming gene expression in multicellular organisms for physiology modulation through engineered bacteria
- Authors: Gao, Baizhen; Sun, Qing
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-22894-7
- Institution: Texas A&M University, USA
- A Computational Framework for Identifying Promoter Sequences in Nonmodel Organisms Using RNA-seq Data Sets
- Authors: Wilson, Erin H; Groom, Joseph D; Sarfatis, M Claire; Ford, Stephanie M; Lidstrom, Mary E; Beck, David A C
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.1c00017
- Institution: University of Washington, USA
Computational Biology / Bioinformatics
Representation Learning
核酸、アミノ酸配列の表現学習- Light Attention Predicts Protein Location from the Language of Life
- Authors: Stärk, Hannes; Dallago, Christian; Heinzinger, Michael; Rost, Burkhard
- Journal: bioRxiv
- Year: 2021
- DOI: 10.1101/2021.04.25.441334
- Institution: Technical University of Munich, Germany
普通のfine-tuningとの比較もして欲しいところ
Other Biology
- A third purine biosynthetic pathway encoded by aminoadenine-based viral DNA genomes
- Authors: Sleiman, Dona; Garcia, Pierre Simon; Lagune, Marion; Loc'h, Jerome; Haouz, Ahmed; Taib, Najwa; Röthlisberger, Pascal; Gribaldo, Simonetta; Marlière, Philippe; Kaminski, Pierre Alexandre
- Journal: Science
- Year: 2021
- DOI: 10.1126/science.abe6494
- Institution: Institut Pasteur, France
合成生物学論文メモ (Mar 2021)
読み流した論文のメモ。黒色はメモ、緑色は感想、赤色は特に面白いと思ったもの。
27報
Synthetic Biology
Transcription/Translation Control
遺伝子回路、転写翻訳制御など- Reversible thermal regulation for bifunctional dynamic control of gene expression in Escherichia coli
- Authors: Wang, Xuan; Han, Jia-Ning; Zhang, Xu; Ma, Yue-Yuan; Lin, Yina; Wang, Huan; Li, Dian-Jie; Zheng, Tao-Ran; Wu, Fu-Qing; Ye, Jian-Wen; Chen, Guo-Qiang
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21654-x
- Institution: Tsinghua University, China
温度センサーだからこそ出来る事があると面白そう。
- Single-cell measurement of plasmid copy number and promoter activity
- Authors: Shao, Bin; Rammohan, Jayan; Anderson, Daniel A; Alperovich, Nina; Ross, David; Voigt, Christopher A
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21734-y
- Institution: MIT, USA
複製開始点の定量的なデザインが出来そう。
RNA Synthetic Biology
アプタマー、リボザイムなど- A multiplexed, automated evolution pipeline enables scalable discovery and characterization of biosensors
- Authors: Townshend, Brent; Xiang, Joy S; Manzanarez, Gabriel; Hayden, Eric J; Smolke, Christina D
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21716-0
- Institution: Stanford University, USA
SELEXの改良版のような感じ、候補の中から当たりを効率的に選べるのが強い。
- Guanidine Biosensors Enable Comparison of Cellular Turn-on Kinetics of Riboswitch-Based Biosensor and Reporter
- Authors: Manna, Sudeshna; Truong, Johnny; Hammond, Ming C
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00583
- Institution: University of Utah, USA
設計の各ステップでの論理展開が明快。
- Overcoming the design, build, test bottleneck for synthesis of nonrepetitive protein-RNA cassettes
- Authors: Katz, Noa; Tripto, Eitamar; Granik, Naor; Goldberg, Sarah; Atar, Orna; Yakhini, Zohar; Orenstein, Yaron; Amit, Roee
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21578-6
- Institution: Israel Institute of Technology, Israel
予測モデルまで組めているので、in silicoデザイン出来ると良いなあ。
Cell-free / Reconstruction
無細胞系、再構成など- A self-organized synthetic morphogenic liposome responds with shape changes to local light cues
- Authors: Gavriljuk, Konstantin; Scocozza, Bruno; Ghasemalizadeh, Farid; Seidel, Hans; Nandan, Akhilesh P; Campos-Medina, Manuel; Schmick, Malte; Koseska, Aneta; Bastiaens, Philippe I H
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21679-2
- Institution: Max Planck Institute, Germany
- Small Antisense DNA-Based Gene Silencing Enables Cell-Free Bacteriophage Manipulation and Genome Replication
- Authors: Vogele, Kilian; Falgenhauer, Elisabeth; von Schönberg, Sophie; Simmel, Friedrich C; Pirzer, Tobias
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00402
- Institution: Technische Universität München, Germany
Optogenetics
光駆動型ツール、蛍光イメージング、光受容体など、その他〇〇genetics系- A Reporter System for Cytosolic Protein Aggregates in Yeast
- Authors: Romero-Suarez, David; Wulff, Tune; Rong, Yixin; Jakočiu Nas, Tadas; Yuzawa, Satoshi; Keasling, Jay D; Jensen, Michael K
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00476
- Institution: Technical University of Denmark, Denmark
オミクス解析もしていて、論理展開が丁寧。
- Optogenetic Modification of Pseudomonas aeruginosa Enables Controllable Twitching Motility and Host Infection
- Authors: Xia, Aiguo; Qian, Mingjie; Wang, Congcong; Huang, Yajia; Liu, Zhi; Ni, Lei; Jin, Fan
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00559
- Institution: Chinese Academy of Sciences, China
- Structural Determinants for Light-Dependent Membrane Binding of a Photoswitchable Polybasic Domain
- Authors: Li, Ling; He, Lian; Wu, Bo; Yu, Chuandi; Zhao, Hongxin; Zhou, Yubin; Wang, Junfeng; Zhu, Lei
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00571
- Institution: Chinese Academy of Sciences, China
- pHmScarlet is a pH-sensitive red fluorescent protein to monitor exocytosis docking and fusion steps
- Authors: Liu, Anyuan; Huang, Xiaoshuai; He, Wenting; Xue, Fudong; Yang, Yanrui; Liu, Jiajia; Chen, Liangyi; Yuan, Lin; Xu, Pingyong
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21666-7
- Institution: University of Science and Technology of China, China
Protein Engineering
タンパク質工学- Random and combinatorial mutagenesis for improved total production of secretory target protein in Escherichia coli
- Authors: Gonzalez-Perez, David; Ratcliffe, James; Tan, Shu Khan; Wong, Mary Chen May; Yee, Yi Pei; Nyabadza, Natsai; Xu, Jian-He; Wong, Tuck Seng; Tee, Kang Lan
- Journal: Scientific Reports
- Year: 2021
- DOI: 10.1038/s41598-021-84859-6
- Institution: University of Sheffield, UK
比較的自由な分泌機構が出来るとQSシグナルのような形で使えそう。
- De novo design of a reversible phosphorylation-dependent switch for membrane targeting
- Authors: Harrington, Leon; Fletcher, Jordan M; Heermann, Tamara; Woolfson, Derek N; Schwille, Petra
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21622-5
- Institution: Max Planck Institute , Germany
coiled-coilをどう変異させるかの理由付けが明快で分かりやすかった。
- Polymerase-guided base editing enables in vivo mutagenesis and rapid protein engineering
- Authors: Cravens, Aaron; Jamil, Osman K; Kong, Deze; Sockolosky, Jonathan T; Smolke, Christina D
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21876-z
- Institution: Stanford University, USA
シチジンデアミナーゼを使っている影響でC>T変異が多いけど、実用上は問題ないとのこと。
CRISPR/Cas
クリスパー系- Programmable C:G to G:C genome editing with CRISPR-Cas9-directed base excision repair proteins
- Authors: Chen, Liwei; Park, Jung Eun; Paa, Peter; Rajakumar, Priscilla D; Prekop, Hong-Ting; Chew, Yi Ting; Manivannan, Swathi N; Chew, Wei Leong
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21559-9
- Institution: Genome Institute of Singapore, Singapore
- Inherently confinable split-drive systems in Drosophila
- Authors: Terradas, Gerard; Buchman, Anna B; Bennett, Jared B; Shriner, Isaiah; Marshall, John M; Akbari, Omar S; Bier, Ethan
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21771-7
- Institution: UCSD, USA
- Inhibition of CRISPR-Cas12a DNA targeting by nucleosomes and chromatin
- Authors: Strohkendl, Isabel; Saifuddin, Fatema A; Gibson, Bryan A; Rosen, Michael K; Russell, Rick; Finkelstein, Ilya J
- Journal: Science Advances
- Year: 2021
- DOI: 10.1126/sciadv.abd6030
- Institution: University of Texas at Austin, USA
Metabolic/Signal Pathway Engineering
シグナル経路、代謝経路、酵素工学など- Metabolic Engineering of Saccharomyces cerevisiae for Ethyl Acetate Biosynthesis
- Authors: Shi, Wenqi; Li, Jie; Chen, Yanfang; Liu, Xiaohang; Chen, Yefu; Guo, Xuewu; Xiao, Dongguang
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00446
- Institution: Tianjin University of Science and Technology, China
DNA / Biophysics
DNA、核酸論理回路、ナノスケール構造物など- Engineering nucleosomes for generating diverse chromatin assemblies
- Authors: Adhireksan, Zenita; Sharma, Deepti; Lee, Phoi Leng; Bao, Qiuye; Padavattan, Sivaraman; Shum, Wayne K; Davey, Gabriela E; Davey, Curt A
- Journal: Nucleic Acids Research
- Year: 2021
- DOI: 10.1093/nar/gkab070
- Institution: Nanyang Technological University, Singapore
Alternative Hosts / Strain Engineering
宿主や系統株の開発- DIVERSIFY: A Fungal Multispecies Gene Expression Platform
- Authors: Jarczynska, Zofia D; Rendsvig, Jakob K H; Pagels, Nichlas; Viana, Veronica R; Nødvig, Christina S; Kirchner, Ferdinand H; Strucko, Tomas; Nielsen, Michael L; Mortensen, Uffe H
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00587
- Institution: Technical University of Denmark, Denmark
Computational Biology / Bioinformatics
Representation Learning
核酸、アミノ酸配列の表現学習- Evotuning protocols for Transformer-based variant effect prediction on multi-domain proteins
- Authors: Yamaguchi, Hideki; Saito, Yutaka
- Journal: bioRxiv
- Year: 2021
- DOI: 10.1101/2021.03.05.434175
- Institution: University of Tokyo, Japan
タスクごとのfine-tuning方法に選択肢を与える研究。ホモログのデータセットをどう集めるかがタスクによっては難しいかも。
Other Biology
- RNA thermoswitches modulate Staphylococcus aureus adaptation to ambient temperatures
- Authors: Catalan-Moreno, Arancha; Cela, Marta; Menendez-Gil, Pilar; Irurzun, Naiara; Caballero, Carlos J; Caldelari, Isabelle; Toledo-Arana, Alejandro
- Journal: Nucleic Acids Research
- Year: 2021
- DOI: 10.1093/nar/gkab117
- Institution: CSIC-Gobierno de Navarra, Spain
そのままRNAスイッチとして移植出来そう。
- Multi-omics Analysis of CRISPRi-Knockdowns Identifies Mechanisms that Buffer Decreases of Enzymes in E. coli Metabolism
- Authors: Donati, Stefano; Kuntz, Michelle; Pahl, Vanessa; Farke, Niklas; Beuter, Dominik; Glatter, Timo; Gomes-Filho, José Vicente; Randau, Lennart; Wang, Chun-Ying; Link, Hannes
- Journal: Cell Systems
- Year: 2021
- DOI: 10.1016/j.cels.2020.10.011
- Institution: Max Planck Institute, Germany
- Binding site profiles and N-terminal minor groove interactions of the master quorum-sensing regulator LuxR enable flexible control of gene activation and repression
- Authors: Zhang, Jun; Liu, Bing; Gu, Dan; Hao, Yuan; Chen, Mo; Ma, Yue; Zhou, Xiaohui; Reverter, David; Zhang, Yuanxing; Wang, Qiyao
- Journal: Nucleic Acids Research
- Year: 2021
- DOI: 10.1093/nar/gkab150
- Institution: East China University of Science and Technology, China
- Parallel molecular mechanisms for enzyme temperature adaptation
- Authors: Pinney, Margaux M; Mokhtari, Daniel A; Akiva, Eyal; Yabukarski, Filip; Sanchez, David M; Liang, Ruibin; Doukov, Tzanko; Martinez, Todd J; Babbitt, Patricia C; Herschlag, Daniel
- Journal: Science
- Year: 2021
- DOI: 10.1126/science.aay2784
- Institution: Stanford University, USA
合成生物学論文メモ (Feb 2021)
読み流した論文のメモ。黒色はメモ、緑色は感想、赤色は特に面白いと思ったもの。
33報
Synthetic Biology
Transcription/Translation Control
遺伝子回路、転写翻訳制御など- Synthetic protein quality control to enhance full-length translation in bacteria
- Authors: Yang, Jina; Han, Yong Hee; Im, Jongwon; Seo, Sang Woo
- Journal: Nature Chemical Biology
- Year: 2021
- DOI: 10.1038/s41589-021-00736-3
- Institution: Seoul National University, Korea
sfGFPだけを発現させた場合はQCシステムと通常のtoehold switchとで発現量は90%ぐらいで済んでいるが、どこまで遺伝子を長く出来るのかは気になるところ。
- Winner-takes-all resource competition redirects cascading cell fate transitions
- Authors: Zhang, Rong; Goetz, Hanah; Melendez-Alvarez, Juan; Li, Jiao; Ding, Tian; Wang, Xiao; Tian, Xiao-Jun
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21125-3
- Institution: Arizona State University, USA
- CRISPRi-Mediated NIMPLY Logic Gate for Fine-Tuning the Whole-Cell Sensing toward Simple Urine Glucose Detection
- Authors: Tan, Shih-I; Ng, I-Son
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.1c00014
- Institution: National Cheng Kung University, China
dCasを使った制御はデザイン空間が広くて最適化が大変そう。
- Potential Landscapes, Bifurcations, and Robustness of Tristable Networks
- Authors: Dey, Anupam; Barik, Debashis
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00570
- Institution: University of Hyderabad, India
wetでの実装も楽しみ。
- Rational Design of the N-Terminal Coding Sequence for Regulating Enzyme Expression in Bacillus subtilis
- Authors: Xu, Kuidong; Tong, Yi; Li, Yi; Tao, Jin; Li, Jianghua; Zhou, Jingwen; Liu, Song
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00309
- Institution: Jiangnan University, China
結構変わるものですね。
- Systematic Quantification of Sequence and Structural Determinants Controlling mRNA stability in Bacterial Operons
- Authors: Cetnar, Daniel P; Salis, Howard M
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00471
- Institution: Pennsylvania State University, USA
他の論文と合わせて大きめの定量的な予測モデルに成り得そう。
- Auxin-mediated protein depletion for metabolic engineering in terpene-producing yeast
- Authors: Lu, Zeyu; Peng, Bingyin; Ebert, Birgitta E; Dumsday, Geoff; Vickers, Claudia E
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21313-1
- Institution: University of Queensland, Australia
ユビキチン化でのタンパク質分解ってどのレベルまで壊すのだろうか。無細胞系で調節可能な環境収容力として使えたりしたら面白そう。
- Model-guided design of mammalian genetic programs
- Authors: Muldoon, J J; Kandula, V; Hong, M; Donahue, P S; Boucher, J D; Bagheri, N; Leonard, J N
- Journal: Science Advances
- Year: 2021
- DOI: 10.1126/sciadv.abe9375
- Institution: Northwestern University, USA
これはシンプルに凄い。COMETシステム=>合成生物学論文メモ(Feb. 2020)
Cell-free / Reconstruction
無細胞系、再構成など- Chromatophores efficiently promote light-driven ATP synthesis and DNA transcription inside hybrid multicompartment artificial cells
- Authors: Altamura, Emiliano; Albanese, Paola; Marotta, Roberto; Milano, Francesco; Fiore, Michele; Trotta, Massimo; Stano, Pasquale; Mavelli, Fabio
- Journal: PNAS
- Year: 2021
- DOI: 10.1073/pnas.2012170118
- Institution: University of Bari Aldo Moro, Italy
合成したATPを使った下流の反応が盛んに出来てくると大分再構成の幅が広がりそう。
- Cell-free Directed Evolution of a Protease in Microdroplets at Ultrahigh Throughput
- Authors: Holstein, Josephin M; Gylstorff, Christian; Hollfelder, Florian
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00538
- Institution: University of Cambridge, UK
ドロップレットを蛍光強度で分別する部分が改善できれば良いが、マイクロ流路かつcell-freeとなるとコスト的にどうなんだという疑問はある。
- A Streptomyces venezuelae Cell-Free Toolkit for Synthetic Biology
- Authors: Moore, Simon J; Lai, Hung-En; Chee, Soo-Mei; Toh, Ming; Coode, Seth; Chengan, Kameshwari; Capel, Patrick; Corre, Christophe; de Los Santos, Emmanuel Lc; Freemont, Paul S
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00581
- Institution: Imperial College London, UK
- Allosteric Regulation of DNA Circuits Enables Minimal and Rapid Biosensors of Small Molecules
- Authors: Rodríguez-Serrano, Alan F; Hsing, I-Ming
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00545
- Institution: Hong Kong University of Science and Technology, Hong Kong
DNAのハイブリダイゼーションを使う事でRNAを介さずにバイオセンサーを作っていて面白い。保存もし易そう。
Optogenetics
光駆動型ツール、蛍光イメージング、光受容体など、その他〇〇genetics系- Dynamical Modeling of Optogenetic Circuits in Yeast for Metabolic Engineering Applications
- Authors: Lovelett, Robert J; Zhao, Evan M; Lalwani, Makoto A; Toettcher, Jared E; Kevrekidis, Ioannis G; L Avalos, José
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00372
- Institution: Princeton University, USA
- Optogenetics in Sinorhizobium meliloti Enables Spatial Control of Exopolysaccharide Production and Biofilm Structure
- Authors: Pirhanov, Azady; Bridges, Charles M; Goodwin, Reed A; Guo, Yi-Syuan; Furrer, Jessica; Shor, Leslie M; Gage, Daniel J; Cho, Yong Ku
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00498
- Institution: University of Connecticut, USA
Protein Engineering
タンパク質工学- Computation-guided optimization of split protein systems
- Authors: Dolberg, Taylor B; Meger, Anthony T; Boucher, Jonathan D; Corcoran, William K; Schauer, Elizabeth E; Prybutok, Alexis N; Raman, Srivatsan; Leonard, Joshua N
- Journal: Nature Chemical Biology
- Year: 2021
- DOI: 10.1038/s41589-020-00729-8
- Institution: Northwestern University, USA
確実に非効率なものは部分的に取り除けているが、まだまだ当たりの予測は難しそう。
- Identification of efficient prokaryotic cell-penetrating peptides with applications in bacterial biotechnology
- Authors: Lee, Hyang-Mi; Ren, Jun; Tran, Kha Mong; Jeon, Byeong-Min; Park, Won-Ung; Kim, Hyunjoo; Lee, Kyung Eun; Oh, Yuna; Choi, Myungback; Kim, Dae-Sung; Na, Dokyun
- Journal: Communications Biology
- Year: 2021
- DOI: 10.1038/s42003-021-01726-w
- Institution: Korea University, Korea
データがもう少し大きければin silicoペプチド設計のターゲットとして向いていそう。
- De novo design of transmembrane β barrels
- Authors: Vorobieva, Anastassia A; White, Paul; Liang, Binyong; Horne, Jim E; Bera, Asim K; Chow, Cameron M; Gerben, Stacey; Marx, Sinduja; Kang, Alex; Stiving, Alyssa Q; Harvey, Sophie R; Marx, Dagan C; Khan, G Nasir; Fleming, Karen G; Wysocki, Vicki H; Brockwell, David J; Tamm, Lukas K; Radford, Sheena E; Baker, David
- Journal: Science
- Year: 2021
- DOI: 10.1126/science.abc8182
- Institution: University of Washington, USA
一つ一つのデザインステップが理論的のどの程度妥当で再現可能なのかよく分からない、解説求む。
CRISPR/Cas
クリスパー系- Programmable human histone phosphorylation and gene activation using a CRISPR/Cas9-based chromatin kinase
- Authors: Li, Jing; Mahata, Barun; Escobar, Mario; Goell, Jacob; Wang, Kaiyuan; Khemka, Pranav; Hilton, Isaac B
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21188-2
- Institution: Rice University, USA
どの制御因子がデザイン上重要かが分かるようになるとより面白い。
- Cas12a-assisted precise targeted cloning using in vivo Cre-lox recombination
- Authors: Enghiad, Behnam; Huang, Chunshuai; Guo, Fang; Jiang, Guangde; Wang, Bin; Tabatabaei, S Kasra; Martin, Teresa A; Zhao, Huimin
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21275-4
- Institution: University of Illinois at Urbana-Champaign, USA
バイオインフォで遺伝子クラスター特定->検証の流れが簡単になりそう。
- Microbial single-strand annealing proteins enable CRISPR gene-editing tools with improved knock-in efficiencies and reduced off-target effects
- Authors: Wang, Chengkun; Cheng, Jason K W; Zhang, Qianhe; Hughes, Nicholas W; Xia, Qiong; Winslow, Monte M; Cong, Le
- Journal: Nucleic Acids Research
- Year: 2021
- DOI: 10.1093/nar/gkaa1264
- Institution: Stanford University, USA
Metabolic/Signal Pathway Engineering
シグナル経路、代謝経路、酵素工学など- A glucose meter interface for point-of-care gene circuit-based diagnostics
- Authors: Amalfitano, Evan; Karlikow, Margot; Norouzi, Masoud; Jaenes, Katariina; Cicek, Seray; Masum, Fahim; Sadat Mousavi, Peivand; Guo, Yuxiu; Tang, Laura; Sydor, Andrew; Ma, Duo; Pearson, Joel D; Trcka, Daniel; Pinette, Mathieu; Ambagala, Aruna; Babiuk, Shawn; Pickering, Bradley; Wrana, Jeff; Bremner, Rod; Mazzulli, Tony; Sinton, David; Brumell, John H; Green, Alexander A; Pardee, Keith
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20639-6
- Institution: University of Toronto, Canada
発想としては面白いが、バックグラウンドの除去にも苦労しているようだし蛍光タンパクの代替として使う意味があるかは不明。
- Engineering advanced logic and distributed computing in human CAR immune cells
- Authors: Cho, Jang Hwan; Okuma, Atsushi; Sofjan, Katri; Lee, Seunghee; Collins, James J; Wong, Wilson W
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21078-7
- Institution: Boston University, USA
やり方はシンプルだけど実際に出来るところが凄い。免疫細胞同士の特異性の高い細胞間コミュニケーションが設計できたりすると面白い。
- Programmable gene regulation for metabolic engineering using decoy transcription factor binding sites
- Authors: Wang, Tiebin; Tague, Nathan; Whelan, Stephen A; Dunlop, Mary J
- Journal: Nucleic Acids Research
- Year: 2021
- DOI: 10.1093/nar/gkaa1234
- Institution: Boston University, USA
- Macrolide Biosensor Optimization through Cellular Substrate Sequestration
- Authors: Miller, Corwin A; Ho, Joanne M; Parks, Sydney E; Bennett, Matthew R
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00572
- Institution: Rice University, USA
QSの性能向上も似たような方法で出来そうだが、既にありそうでもある。相分離的な隔離とも相性が良さそう。
DNA / Biophysics
DNA、核酸論理回路、ナノスケール構造物など- Self-assembly and regulation of protein cages from pre-organised coiled-coil modules
- Authors: Lapenta, Fabio; Aupič, Jana; Vezzoli, Marco; Strmšek, Žiga; Da Vela, Stefano; Svergun, Dmitri I; Carazo, José María; Melero, Roberto; Jerala, Roman
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21184-6
- Institution: National Institute of Chemistry, Slovenia
タンパク質折り紙もDNAみたいになってきて面白い。タンパク質の方が特異性も高く他のタンパク質との相互作用を組み込めて、色々出来そう。
Alternative Hosts / Strain Engineering
宿主や系統株の開発- Construction of intracellular asymmetry and asymmetric division in Escherichia coli
- Authors: Lin, Da-Wei; Liu, Yang; Lee, Yue-Qi; Yang, Po-Jiun; Ho, Chia-Tse; Hong, Jui-Chung; Hsiao, Jye-Chian; Liao, Der-Chien; Liang, An-Jou; Hung, Tzu-Chiao; Chen, Yu-Chuan; Tu, Hsiung-Lin; Hsu, Chao-Ping; Huang, Hsiao-Chun
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-21135-1
- Institution: National Taiwan University, Taiwan
他の制御因子と組み合わせて複数段階で分化が起こせたりしたら面白そう。
Computational Biology / Bioinformatics
Representation Learning
核酸、アミノ酸配列の表現学習- A transformer architecture based on BERT and 2D convolutional neural network to identify DNA enhancers from sequence information
- Authors: Le, Nguyen Quoc Khanh; Ho, Quang-Thai; Nguyen, Trinh-Trung-Duong; Ou, Yu-Yen
- Journal: Briefings in Bioinformatics
- Year: 2021
- DOI: 10.1093/bib/bbab005
- Institution: Taipei Medical University, Taiwan
CLSトークンを使わずに分類器にかけている辺りBERTだけでは上手くいかなかったっぽいが、単純な分散表現学習器として使ってしまうのは勿体無い気も。
- Accurate deep learning off-target prediction with novel sgRNA-DNA sequence encoding in CRISPR-Cas9 gene editing
- Authors: Charlier, Jeremy; Nadon, Robert; Makarenkov, Vladimir
- Journal: Bioinformatics
- Year: 2021
- DOI: 10.1093/bioinformatics/btab112
- Institution: Université du Québec à Montréal, Canada
- lncLocator 2.0: a cell-line-specific subcellular localization predictor for long non-coding RNAs with interpretable deep learning
- Authors: Lin, Yang; Pan, Xiaoyong; Shen, Hong-Bin
- Journal: Bioinformatics
- Year: 2021
- DOI: 10.1093/bioinformatics/btab127
- Institution: Shanghai Jiao Tong University, China
合成生物学論文メモ (Jan 2021)
読み流した論文のメモ。黒色はメモ、緑色は感想、赤色は特に面白いと思ったもの。
27報
Synthetic Biology
Transcription/Translation Control
遺伝子回路、転写翻訳制御など- Orthogonal control of mean and variability of endogenous genes in a human cell line
- Authors: Bonny, Alain R; Fonseca, João Pedro; Park, Jesslyn E; El-Samad, Hana
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20467-8
- Institution: University of California, San Francisco, USA
発現量の平均と分散をコントロールする発想があったか。
- Multiplexed characterization of rationally designed promoter architectures deconstructs combinatorial logic for IPTG-inducible systems
- Authors: Yu, Timothy C; Liu, Winnie L; Brinck, Marcia S; Davis, Jessica E; Shek, Jeremy; Bower, Grace; Einav, Tal; Insigne, Kimberly D; Phillips, Rob; Kosuri, Sriram; Urtecho, Guillaume
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20094-3
- Institution: UCLA, USA
地味だけど良い研究。
- Fine-Tuning Multi-Gene Clusters via Well-Characterized Gene Expression Regulatory Elements: Case Study of the Arginine Synthesis Pathway in C. glutamicum
- Authors: Duan, Yanting; Zhai, Weiji; Liu, Weijia; Zhang, Xiaomei; Shi, Jin-Song; Zhang, Xiaojuan; Xu, Zhenghong
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00405
- Institution: Jiangnan University, China
bcRBSというのを知らなかったけど、割と便利そう。
- Optimization of a p‐Coumaric Acid Biosensor System for Versatile Dynamic Performance
- Authors: Jiang, Tian; Li, Chenyi; Yan, Yajun
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: DOI
- Institution: The University of Georgia, USA
- Contextual dependencies expand the re-usability of genetic inverters
- Authors: Tas, Huseyin; Grozinger, Lewis; Stoof, Ruud; de Lorenzo, Victor; Goñi-Moreno, Ángel
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20656-5
- Institution: Campus de Cantoblanco, Spain
結構な違いが出ていて驚き。
- Nanobody-mediated control of gene expression and epigenetic memory
- Authors: Van, Mike V; Fujimori, Taihei; Bintu, Lacramioara
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20757-1
- Institution: Stanford University, USA
エピジェネティック工学も回路が組めるようになってきた。
RNA Synthetic Biology
アプタマー、リボザイムなど- Orthogonal translation enables heterologous ribosome engineering in E. coli
- Authors: Kolber, Natalie S; Fattal, Ranan; Bratulic, Sinisa; Carver, Gavriela D; Badran, Ahmed H
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20759-z
- Institution: Broad Institute of MIT and Harvard, USA
Cell-free / Reconstruction
無細胞系、再構成など- In vitro synthesis of 32 translation-factor proteins from a single template reveals impaired ribosomal processivity
- Authors: Doerr, Anne; Foschepoth, David; Forster, Anthony C; Danelon, Christophe
- Journal: Scientific Reports
- Year: 2021
- DOI: 10.1038/s41598-020-80827-8
- Institution: Delft University of Technology, Netherlands
自作のPUREだとやはりバイアスが結構かかってしまうらしい。
- Posttranslational chemical installation of azoles into translated peptides
- Authors: Tsutsumi, Haruka; Kuroda, Tomohiro; Kimura, Hiroyuki; Goto, Yuki; Suga, Hiroaki
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-20992-0
- Institution: The University of Tokyo, Japan
- Site-specific incorporation of citrulline into proteins in mammalian cells
- Authors: Mondal, Santanu; Wang, Shu; Zheng, Yunan; Sen, Sudeshna; Chatterjee, Abhishek; Thompson, Paul R
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20279-w
- Institution: UMass Medical School, USA
Citrullination自体も初めて知ったけど、aaRSの変異で部位特異的に翻訳後修飾を入れられるって凄いな。
Optogenetics
光駆動型ツール、蛍光イメージング、光受容体など、その他〇〇genetics系- Liquid-liquid phase separation of light-inducible transcription factors increases transcription activation in mammalian cells and mice
- Authors: Schneider, Nils; Wieland, Franz-Georg; Kong, Deqiang; Fischer, Alexandra A M; Hörner, Maximilian; Timmer, Jens; Ye, Haifeng; Weber, Wilfried
- Journal: Science Advances
- Year: 2021
- DOI: 10.1126/sciadv.abd3568
- Institution: University of Freiburg, Germany
相分離による転写活性化で、青色光なのに生体マウスの組織深部での転写発現制御が出来ていて強力。相分離の合成生物学はこれから色々出て来そう。
- Rational Design of a Split Flavin-Based Fluorescent Reporter
- Authors: Yudenko, Anna; Smolentseva, Anastasia; Maslov, Ivan; Semenov, Oleg; Goncharov, Ivan M; Nazarenko, Vera V; Maliar, Nina L; Borshchevskiy, Valentin; Gordeliy, Valentin; Remeeva, Alina; Gushchin, Ivan
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00454
- Institution: Moscow Institute of Physics and Technology, Russia
理論的なタンパク質デザインをしていて、非常に良い論文。
- Engineering an Optogenetic CRISPRi Platform for Improved Chemical Production
- Authors: Wu, Peiling; Chen, Yufen; Liu, Mingyu; Xiao, Gezhi; Yuan, Jifeng
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00488
- Institution: Xiamen University, China
Fig3は青色光照射無しとの比較がないとコントロールが取れていない気がする。
- A synthetic BRET-based optogenetic device for pulsatile transgene expression enabling glucose homeostasis in mice
- Authors: Li, Ting; Chen, Xianjun; Qian, Yajie; Shao, Jiawei; Li, Xie; Liu, Shuning; Zhu, Linyong; Zhao, Yuzheng; Ye, Haifeng; Yang, Yi
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-021-20913-1
- Institution: East China University of Science and Technology, China
何故わざわざBRETなのかと思ったら、意外な性能だった。
Protein Engineering
タンパク質工学- Bottom-up de novo design of functional proteins with complex structural features
- Authors: Yang, Che and Sesterhenn, Fabian and Bonet, Jaume and van Aalen, Eva A and Scheller, Leo and Abriata, Luciano A and Cramer, Johannes T and Wen, Xiaolin and Rosset, St{\'e}phane and Georgeon, Sandrine and Jardetzky, Theodore and Krey, Thomas and Fussenegger, Martin and Merkx, Maarten and Correia, Bruno E
- Journal: Nature Chemical Biology
- Year: 2021
- DOI: 10.1038/s41589-020-00699-x. (École Polytechnique Fédérale de Lausann
- Institution: École Polytechnique Fédérale de Lausanne (EPFL), Switzerland
タンパク質デザインあるある: こんな簡単に出来たら苦労しない。
- YESS 2.0, a Tunable Platform for Enzyme Evolution, Yields Highly Active TEV Protease Variants
- Authors: Denard, Carl A; Paresi, Chelsea; Yaghi, Rasha; McGinnis, Natalie; Bennett, Zachary; Yi, Li; Georgiou, George; Iverson, Brent L
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00452
- Institution: University of Texas Austin, USA
- Direct control of CAR T cells through small molecule-regulated antibodies
- Authors: Park, Spencer; Pascua, Edward; Lindquist, Kevin C; Kimberlin, Christopher; Deng, Xiaodi; Mak, Yvonne S L; Melton, Zea; Johnson, Theodore O; Lin, Regina; Boldajipour, Bijan; Abraham, Robert T; Pons, Jaume; Sasu, Barbra Johnson; Van Blarcom, Thomas J; Chaparro-Riggers, Javier
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20671-6
- Institution: Pfizer, USA
CRISPR/Cas
クリスパー系- Engineered dual selection for directed evolution of SpCas9 PAM specificity
- Authors: Goldberg, Gregory W; Spencer, Jeffrey M; Giganti, David O; Camellato, Brendan R; Agmon, Neta; Ichikawa, David M; Boeke, Jef D; Noyes, Marcus B
- Journal: Nature Communications
- Year: 2021
- DOI: 10.1038/s41467-020-20650-x
- Institution: New York University, USA
Metabolic/Signal Pathway Engineering
シグナル経路、代謝経路、酵素工学など- A new-to-nature carboxylation module to improve natural and synthetic CO 2 fixation
- Authors: Scheffen, Marieke and Marchal, Daniel G and Beneyton, Thomas and Schuller, Sandra K and Klose, Melanie and Diehl, Christoph and Lehmann, Jessica and Pfister, Pascal and Carrillo, Martina and He, Hai and Aslan, Selcuk and Cortina, Nina S and Claus, Peter and Bollschweiler, Daniel and Baret, Jean-Christophe and Schuller, Jan M and Zarzycki, Jan and Bar-Even, Arren and Erb, Tobias J
- Journal: Nature Catalysis
- Year: 2021
- DOI: 10.1038/s41929-020-00557-y
- Institution: Max Planck Institute for Terrestrial Microbiology, Germany
酵素活性部位の予測から炭素固定能の計算まで行っていて良い。
- Living materials with programmable functionalities grown from engineered microbial co-cultures
- Authors: Gilbert, Charlie; Tang, Tzu-Chieh; Ott, Wolfgang; Dorr, Brandon A; Shaw, William M; Sun, George L; Lu, Timothy K; Ellis, Tom
- Journal: Nature Materials
- Year: 2021
- DOI: 10.1038/s41563-020-00857-5
- Institution: Imperial College London, UK
共生系の経路の開発は生育/誘導条件の最適化が中々難しそうだが、数理的なデザインはどの程度出来るのかな。
DNA / Biophysics
DNA、核酸論理回路、ナノスケール構造物など- Robust direct digital-to-biological data storage in living cells
- Authors: Yim, Sung Sun; McBee, Ross M; Song, Alan M; Huang, Yiming; Sheth, Ravi U; Wang, Harris H
- Journal: Nature Chemical Biology
- Year: 2021
- DOI: 10.1038/s41589-020-00711-4
- Institution: Columbia University, USA
Alternative Hosts / Strain Engineering
宿主や系統株の開発- CRISPR-based transcriptional activation tool for silent genes in filamentous fungi
- Authors: Mózsik, László; Hoekzema, Mirthe; de Kok, Niels A W; Bovenberg, Roel A L; Nygård, Yvonne; Driessen, Arnold J M
- Journal: Scientific Reports
- Year: 2021
- DOI: 10.1038/s41598-020-80864-3
- Institution: University of Groningen, Netherlands
- Engineering lithoheterotrophy in an obligate chemolithoautotrophic Fe(II) oxidizing bacterium
- Authors: Jain, Abhiney; Gralnick, Jeffrey A
- Journal: Scientific Reports
- Year: 2021
- DOI: 10.1038/s41598-021-81412-3
- Institution: University of Minnesota, USA
Viral Engineering
ファージ関連- Engineering Bacteria to Produce Pure Phage-like Particles for Gene Delivery
- Authors: Tridgett, Matthew; Ababi, Maria; Osgerby, Alexander; Ramirez Garcia, Robert; Jaramillo, Alfonso
- Journal: ACS Synthetic Biology
- Year: 2021
- DOI: 10.1021/acssynbio.0c00467
- Institution: The University of Warwick, France
- Characterizing the portability of phage-encoded homologous recombination proteins
- Authors: Filsinger, Gabriel T; Wannier, Timothy M; Pedersen, Felix B; Lutz, Isaac D; Zhang, Julie; Stork, Devon A; Debnath, Anik; Gozzi, Kevin; Kuchwara, Helene; Volf, Verena; Wang, Stan; Rios, Xavier; Gregg, Christopher J; Lajoie, Marc J; Shipman, Seth L; Aach, John; Laub, Michael T; Church, George M
- Journal: Nature Chemical Biology
- Year: 2021
- DOI: 10.1038/s41589-020-00710-5
- Institution: Harvard University, USA
古典的なバイオっぽい解析と合成生物学的な応用が両方あって良い。
- aeruginosa synthetic phages with reduced genomes
- Authors: Pires, Diana P; Monteiro, Rodrigo; Mil-Homens, Dalila; Fialho, Arsénio; Lu, Timothy K; Azeredo, Joana. Designing P
- Journal: Scientific Reports
- Year: 2021
- DOI: 10.1038/s41598-021-81580-2
- Institution: Universidade Do Minho, Portugal
ウイルスぐらいのゲノムサイズなら予測で同じような事が出来たりしないだろうか。
合成生物学論文メモ (Dec 2020)
読み流した論文のメモ。黒色はメモ、緑色は感想、赤色は特に面白いと思ったもの。
19報
Synthetic Biology
Transcription/Translation Control
遺伝子回路、転写翻訳制御など- Exchange of endogenous and heterogeneous yeast terminators in Pichia pastoris to tune mRNA stability and gene expression
- Authors: Yoichiro Ito, Goro Terai, Misa Ishigami, Noriko Hashiba, Yasuyuki Nakamura, Takahiro Bamba, Ryota Kumokita, Tomohisa Hasunuma, Kiyoshi Asai, Jun Ishii, Akihiko Kondo
- Journal: Nucleic Acids Research
- Year: 2020
- DOI: 10.1093/nar/gkaa1066
- Institution: Kobe University, Japan
- Single Cell Characterization of a Synthetic Bacterial Clock with a Hybrid Feedback Loop Containing dCas9-sgRNA
- Authors: John Henningsen, Matthaeus Schwarz-Schilling, Andreas Leibl, Joaquı́n Gutiérrez, Sandra Sagredo, Friedrich C. Simmel
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00438
- Institution: Technical University of Munich, Germany
位相の性質を実験で調べているのが面白い。
RNA Synthetic Biology
アプタマー、リボザイムなど- Discovery and characterization of a fourth class of guanidine riboswitches
- Authors: Felina Lenkeit, Iris Eckert, Jörg S Hartig, Zasha Weinberg
- Journal: Nucleic Acids Research
- Year: 2020
- DOI: DOI
- Institution: University of Konstanz, Germany
まだこの方法でも良い感じにリボスイッチが見つかるのね。
Cell-free / Reconstruction
無細胞系、再構成などOptogenetics
光駆動型ツール、蛍光イメージング、光受容体など、その他〇〇genetics系- Resurrecting the ancient glow ofthe fireflies
- Authors: Y. Oba, K. Konishi, D. Yano, H. Shibata, D. Kato, T. Shirai
- Journal: Science Advances
- Year: 2020
- DOI: 10.1126/sciadv.abc5705
- Institution: Chubu University, Japan
ものすごく格好いい研究。
- Living materials fabricated via gradient mineralization of light-inducible biofilms
- Authors: Yanyi Wang, Bolin An, Bin Xue, Jiahua Pu, Xiuli Zhang, Yuanyuan Huang, Yi Yu, Yi Cao, Chao Zhong
- Journal: Nature Chemical Biology
- Year: 2020
- DOI: 10.1038/s41589-020-00697-z
- Institution: Shanghai Tech University, China
光強度と厚さで相関が取れているのが凄い。
- Design and Characterization of Rapid Optogenetic Circuits for Dynamic Control in Yeast Metabolic Engineering
- Authors: Evan M. Zhao, Makoto A. Lalwani, Robert J. Lovelett, Sergio A. García-Echauri, Shannon M. Hoffman, Christopher L. Gonzalez, Jared E. Toettcher, Ioannis G. Kevrekidis, José L. Avalos
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00305
- Institution: Princeton University, USA
既存のTFツールとの比較を行なっているところが好感度高い。
- Creating Red Light-Switchable Protein Dimerization Systems as Genetically Encoded Actuators with High Specificity
- Authors: Zhimin Huang, Zengpeng Li, Xiao Zhang, Shoukai Kang, Runze Dong, Li Sun, Xiaonan Fu, David Vaisar, Kurumi Watanabe, Liangcai Gu
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00397
- Institution: University of Washington, Seatle, USA
近赤外光ツールの新作。nanobodyを使って、NIR-light受容体の応答の弱さをカバーしていて、細胞内在性のリガンド(bilverdin)のみで十分な応答が得られているのが強み。ただ、nanobodyを使っているので観察しようと思うと結局別の蛍光タンパクが必要。
Protein Engineering
タンパク質工学- Self-assembly–based posttranslational protein oscillators
- Authors: Ofer Kimchi, Carl P. Goodrich, Alexis Courbet, Agnese I. Curatolo, Nicholas B. Woodall, David Baker, Michael P. Brenner
- Journal: Science Advances
- Year: 2020
- DOI: 10.1126/sciadv.abc1939
- Institution: Harvard University, University of Washington Seatle, USA
二因子の振動が出来たから次は三因子かな。
- In Vivo Biogenesis of a De Novo Designed Iron−Sulfur Protein
- Authors: Bhanu P. Jagilinki, Stefan Ilic, Cristian Trncik, Alexei M. Tyryshkin, Douglas H. Pike, Wolfgang Lubitz, Eckhard Bill, Oliver Einsle, James A. Birrell, Barak Akabayov, Dror Noy, Vikas Nanda
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00514
- Institution: Rutgers University, USA
最小単位の同定まで行なっていて、まとまりのある綺麗な論文。
CRISPR/Cas
クリスパー系- Novel Prokaryotic CRISPR-Cas12a-Based Tool for Programmable Transcriptional Activation and Repression
- Authors: Christoph Schilling, Mattheos A. G. Koffas, Volker Sieber, and Jochen Schmid
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00424
- Institution: Technical University of Munich, Germany
回路素子としての使い所が割とありそう。
- In vivo diversification of target genomic sites using processive base deaminase fusions blocked by dCas9
- Authors: Beatriz Álvarez, Mario Mencía, Víctor de Lorenzo, Luis Ángel Fernández
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-20230-z. (Consejo Superior de Investigaciones Científica
- Institution: Consejo Superior de Investigaciones Científicas (CNB-CSIC), Spain
in vivoで部位特異的&部分的なランダム変異が入れられるのが強み。デアミナーゼのバリエーションや変異の偏りに改善の余地ありといった感じ。
Metabolic/Signal Pathway Engineering
シグナル経路、代謝経路、酵素工学など- Rewiring Central Carbon Metabolism Ensures Increased Provision of Acetyl-CoA and NADPH Required for 3‐OH-Propionic Acid Production
- Authors: Ning Qin, Lingyun Li, Xu Ji, Xiaowei Li, Yiming Zhang, Christer Larsson, Yun Chen, Jens Nielsen, Zihe Liu
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00264
- Institution: Beijing University of Chemical Technology, China
- Efficient Biosynthesis of (2S)‐Eriodictyol from (2S)‐Naringenin in Saccharomyces cerevisiae through a Combination of Promoter Adjustment and Directed Evolution
- Authors: Song Gao, Xiaoyu Xu, Weizhu Zeng, Sha Xu, Yunbin Lyv, Yue Feng, Guoyin Kai, Jingwen Zhou, Jian Chen
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00346
- Institution: Jiangnan University, China
DNA / Biophysics
DNA、核酸論理回路、ナノスケール構造物など- Characterizing and Controlling Nanoscale Self-Assembly of Suckerin-12
- Authors: Jasmine M. Hershewe, William D. Wiseman, James E. Kath, Chelsea C. Buck, Maneesh K. Gupta, Patrick B. Dennis, Rajesh R. Naik, Michael C. Jewett
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00442
- Institution: Northwestern University, USA
Biochemistry
生化学系Alternative Hosts / Strain Engineering
宿主や系統株の開発- Fine-Tuning of Transcription in Pichia pastoris Using dCas9 and RNA Scaffolds
- Authors: Michael Baumschabl, Roland Prielhofer, Diethard Mattanovich, and Matthias G. Steiger
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c0021
- Institution: Austrian Centre of Industrial Biotechnology, Austria
P.pastorisはメタノールの代謝系で強いらしい、ターミネーターの論文でも使われていた株。
- A Logic NAND Gate for Controlling Gene Expression in a Circadian Rhythm in Cyanobacteria
- Authors: Mieun Lee, Han Min Woo
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00455
- Institution: Sungkyunkwan University, Korea
シアノバクテリアだとこういう光応答性の使い方が出来るところのは面白い。
- Automating Cloning by Natural Transformation
- Authors: Xinglin Jiang, Emilia Palazzotto, Ewa Wybraniec, Lachlan Jake Munro, Haibo Zhang, Douglas B. Kell, Tilmann Weber, Sang Yup Lee
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00240
- Institution: Technical University of Denmark, Denmark
既存の形質転換手法を置き換えるほどのメリットは無さそうだけど、この技術を元に何かは出来そう。
細菌に出来事の数え方を教えるお話
Advent Calender「今年読んだ一番好きな論文2020」の記事です。
3行で紹介
2020年9月にNature Communicationsに出たこちらの論文を紹介します。
細菌の群れが出来事の種類と回数を記憶出来る仕組みを、人工的に効率よく作れるようにしましたというお話です。出来事の種類と回数に応じてDNAが書き換えられるようなシステムを構築し、遺伝子から発現する蛍光タンパク質の色を変えています。
生物の仕組みをつくる研究の流れ
今回の紹介論文は、仕組みをつくる流れの研究の一つで、細菌の遺伝情報の中に論理演算の仕組みを組み込んで、ヒトがデザインした通りに出来事を数えさせようというものです。
数を数えさせる仕組み
細菌に数を数えさせる仕組みの鍵となるのは、インテグラーゼ(integrase)と呼ばれる酵素です。この酵素は、DNA上の2種類のマーカー配列(attB, attP)を認識して、そのマーカーの方向によってDNAに2つの異なった作用を不可逆的に施します。
- マーカーが向き合っている場合: 間のDNAを反転させる(画像左側の反応)
- マーカーが同一方向の場合: 間のDNAを切り取る(画像右側の反応)
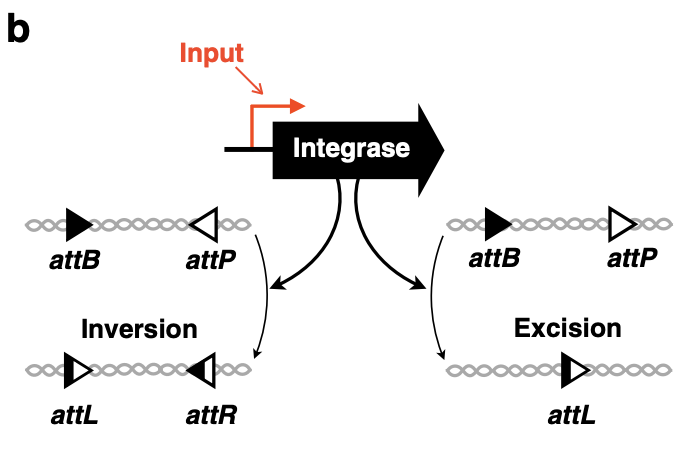
DNAマーカーが向き合っていれば反転、同一方向なら切り出しが起こる。
このようなインテグラーゼを2種類組み合わせると、aとbの2種類の出来事(薬剤等による刺激)の順番によって別々の挙動を示す回路が出来上がります。下図の太い矢印(0, 1, 2)はそれぞれの色の蛍光タンパク質がコードされた遺伝子を表していて、遺伝子は方向性をもつため特定の方向でコードされている場合のみ正常にはたらきます。下図の曲がった矢印は、プロモーターと呼ばれるDNA領域で、遺伝子が読み取られる方向を決定しています。なお、プロモーター自体は左右どちらの方向であっても正常に機能することが出来ます。出来事が起こる前(0の状態)では、青色のタンパク質が発現していて、出来事の種類によって2種類のインテグラーゼのどちらかがはたらき、先に述べたDNAの反転または切り出しが起こります。
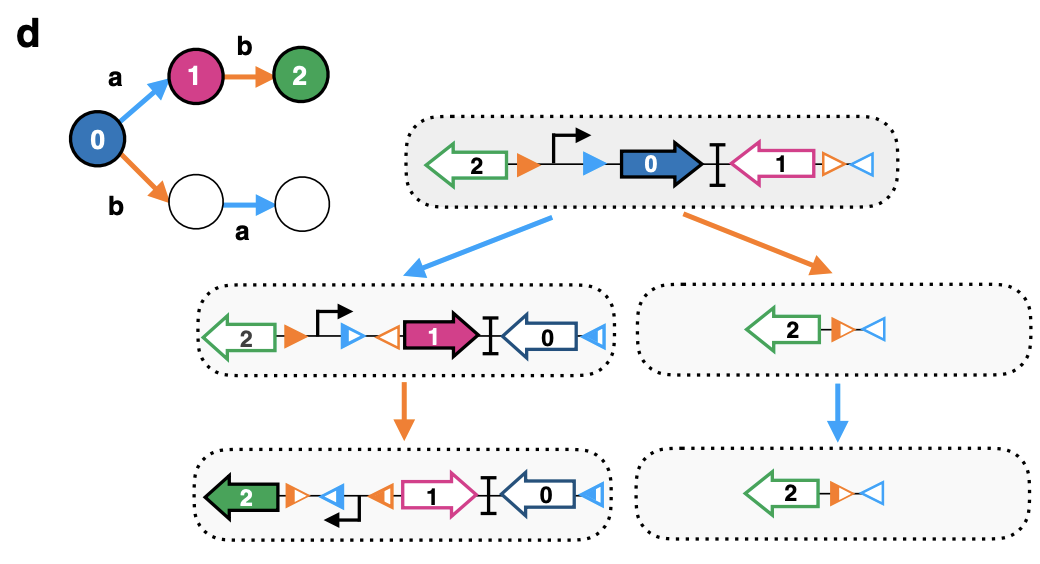
a→bの順番なら、青色→赤色→緑色の順に発現する遺伝子が変化し、b→の順番なら、青色→無→無となる。
- 出来事b(右側、オレンジ色の矢印)が起こった場合
オレンジ色のマーカーにインテグラーゼが働き、マーカーは同一方向のためDNAが切り出されます。正しい方向の蛍光タンパク質もまだ使われていない水色のマーカーも無くなってしまいます。
- 出来事a(左側、水色の矢印)が起こった場合
水色のマーカーにインテグラーゼがはたらき、マーカーが向き合っているのでDNAが反転します。結果として、青色のタンパク質は方向が逆になってしまって機能を失い、代わりに正しい方向になった赤色のタンパク質が発現します。また、オレンジ色のマーカーが向き合うため、引き続いて出来事bが起こると、オレンジ色のマーカーの間でDNAの反転が起こり、プロモーターが逆向きになります。これによって遺伝子の読み取り方向がプロモーターから左側に進ようになるため、緑色のタンパク質が代わりに発現するようになります。
このようにして、細菌が出来事の数と順番を数え、それに応じて異なる遺伝子を発現させる仕組みが出来上がりました。ちなみに、ここまでの研究は2013年に発表されたもの*5の追試です。
より複雑な出来事の数え方
前節までの話は、すでに先行研究で発表されていた内容でした。紹介論文では、細菌がより複雑な出来事も正確に数えられるように仕組みを発展させ、また、出来事の順番を好きなように入れ替えても上手くはたらくようにデザインの効率化を行いました。
その1: 複数の細菌株に分ける
複雑化のその1では、それまで一種類の細菌株で行なっていた演算を複数の株に分けるシステムを実装しました。このシステムの主目的は、演算がより複雑になった場合に、一つの細菌では演算が処理仕切れなくなってしまう事態(具体的には、遺伝子の発現量の限界や導入する遺伝子の合計サイズの限界)を防ぐ事や、出力部分でより複雑な反応をさせたい場合に、反応同士が混み合ってしまう事態を防ぐ事などにあります。
拡張したシステムの生物学的な実装の方法としては、一種類の細菌が保持していた遺伝子を複数の細菌に分けてあげました。例として下図3cを見てみると、青色と緑色蛍光タンパク質の部分を2SP8株に、赤色蛍光タンパク質の部分を2SP5株に分配しています。
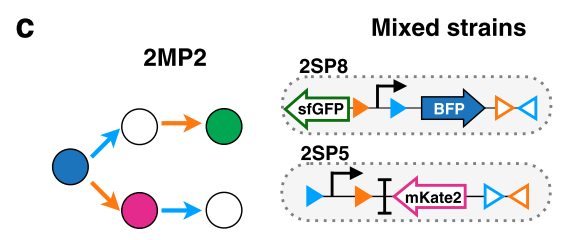
その2: 数えられる出来事の種類を増やす
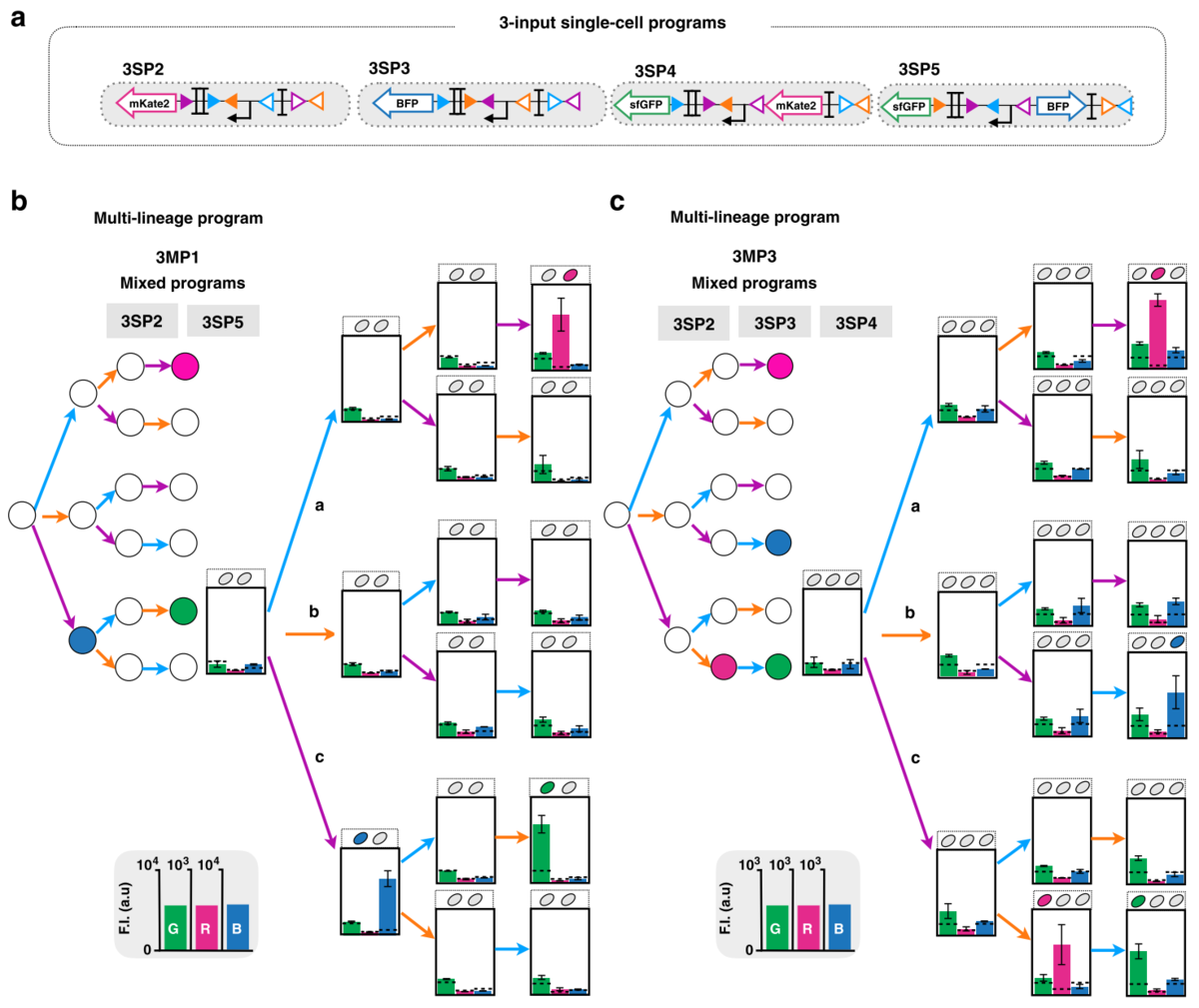
複雑化のその2では、数えられる出来事の種類を増やして演算の自由度を上げました。これによって、現実的に演算を複雑化させる事が可能になります。
以下の図4cには、3種類の出来事を数えられる細菌の例が示されています。この例では、出来事がc→b→aの順番で起こった場合のみ蛍光タンパク質の色の変化が見られます。

複雑な系の実装
複雑化のその1とその2を組み合わせて出来たシステムの様子と結果(棒グラフ)が以下の図6に示されています。aに示されている細菌株の組み合わせによって、出来事の数え方が異なっている様子がbとcとで比較されています。
まとめ
仕組みの実装をするという事を結構簡単に書いてしまいましたが、実際の実験では予期しないところで蛍光タンパク質が発現してしまったり(図6cでは少し残っている)、発現しているはずの蛍光タンパク質が見られなかったりする事が多々(多々)あるので、結構泥臭い最適化が必要だったりします。省略しましたが図で示されている回路のデザインは筆者らが主張する効率の良い最適化スキームで作られていて、この仕組みを応用に使う研究者や企業が他のインテグラーゼや細菌で検証を行う事が期待されます。
今回の話を面白いと思った方は是非↓の論文やレビューも参照してみてください。
合成生物学論文メモ (Nov 2020)
読み流した論文のメモ。黒色はメモ、緑色は感想、赤色は特に面白いと思ったもの。
28報
Synthetic Biology
Transcription/Translation Control
遺伝子回路、転写翻訳制御など- Interpretation of morphogen gradients by a synthetic bistable circuit
- Authors: P.K. Grant, G. Szep, O. Patange, J. Halatek, V. Coppard, A. Csikász-Nagy, J. Haseloff, J.C.W. Locke, N. Dalchau & A. Phillips
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19098-w
- Institution: Microsoft Research, UK
応用へのアプローチの仕方が良い。
- An endoribonuclease-based feedforward controller for decoupling resource-limited genetic modules in mammalian cells
- Authors: R.D. Jones, Y. Qian, V. Siciliano, B. DiAndreth, J. Huh, R. Weiss & D.D. Vecchio
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19126-9
- Institution: MIT, USA
Zhao labと同じmRNA分解による細胞内資源再分配だが、使用している回路の構成が違って面白い。こっちの方が数理モデルからの厳密な定量がされているっぽい?
- Predictive design of sigma factor-specific promoters
- Authors: M.V. Brempt, J. Clauwaert, F. Mey, M. Stock, J. Maertens, W. Waegeman & M.D. Mey
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19446-w
- Institution: Ghent University, Belgium
酵母プロモーター予測器のE.coli版。
- Tunable Gene Expression System Independent of Downstream Coding Sequence
- Authors: S.M. Yoo, S.-W. Jung, J. Yeom, S.Y. Lee & D. Na
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00029
- Institution: Chung-Ang University, Korea
UTRの編集はアイデアとして他にもありそうだけど、どうだろう
- Multidimensional Characterization of Parts Enhances Modeling Accuracy in Genetic Circuits
- Authors: M. Gómez-Schiavon, G. Dods, H. El-Samad, A.H. Ng
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00288
- Institution: UCSF, USA
fitting出来ているものはあるけど、出来ていないものもちらほら。汎用モデルを立てるのはなかなか難しそう。
- Design of Multipartite Transcription Factors for Multiplexed Logic Genome Integration Control in Mammalian Cells
- Authors: S. Ausländer, D. Ausländer, P.F. Lang, M. Kemi & M. Fussenegger
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00413
- Institution: ETH Zurich, Switzerland
人工TFの作り方論文が哺乳類細胞まで来た。
- Synthetic protein-binding DNA sponge as a tool to tune gene expression and mitigate protein toxicity
- Authors: X. Wan, F. Pinto, L. Yu, B. Wang
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19552-9
- Institution: University of Edinburgh, UK
何で今まで誰もやらなかったのかが疑問なレベルのシンプルな発想ながら、強力な結果。
RNA Synthetic Biology
アプタマー、リボザイムなど- Aptamer-Mediated Control of Polyadenylation for Gene Expression Regulation in Mammalian Cells
- Authors: M. Spöring, R. Boneberg & J.S. Hartig
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00222
- Institution: University of Konstanz, Germany
ポリAシグナルの阻害とはその手があったか。3'UTRのリボスイッチの可能性も広げる。
Cell-free / Reconstruction
無細胞系、再構成など- Microcompartmentalized Cell-Free Protein Synthesis in Hydrogel μ‑Channels
- Authors: A.I. Benítez-Mateos, N. Zeballos, N. Comino, L.M. de Redrojo, T. Randelovic, & F. López-Galle*
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00462
- Institution: CICbiomaGUNE, Spain
- From deterministic to fuzzy decision-making in artificial cells
- Authors: F. Greiss, S.S. Daube, V. Noireaux & R. Bar-Ziv
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19395-4
- Institution: Weizmann Institute of Scienc, Israel
キャピラリーで区切っただけで膜のない空間をartificial cellと呼ぶのかは疑問。
- Decoupling Protein Production from Cell Growth Enhances the Site- Specific Incorporation of Noncanonical Amino Acids in E. coli
- Authors: M.G. Casas, P. Stargardt, J. Mairhofer & B. Wiltschi
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00298
- Institution: Austrian Center of Industrial Biotechnology, Austria
非天然アミノ酸もいよいよ現実的なタンパク質デザインが出て来そう。
Optogenetics
光駆動型ツール、蛍光イメージング、光受容体など、その他〇〇genetics系- Optogenetic Control of the BMP Signaling Pathway
- Authors: P.A. Humphreys, S. Woods, C.A. Smith, N. Bates, S.A. Cain, R. Lucas & S.J. Kimber
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00315
- Institution: The University of Manchester, UK
characterization(fig2)が丁寧で良い。
- Genetically Encoded Fluorescent Biosensor for Rapid Detection of Protein Expression
- Authors: M.G. Eason, A.T. Pandelieva, M.M. Mayer, S.T. Khan, H.G. Garcia, R.A. Chica
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00407
- Institution: University of Ottawa, Canada
リアルタイム観察が出来るような応答の速さがポイント高いらしい。
Protein Engineering
タンパク質工学- Scalable continuous evolution for the generation of diverse enzyme variants encompassing promiscuous activities
- Authors: G. Rix, E.J. Watkins-Dulaney, P.J. Almhjell, C.E. Boville, F.H. Arnold & C.C. Liu
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19539-6
- Institution: UC Irvine, USA
eVOLVERでやったらもっと簡単に実現できそう。continuous directed evolutionやりたい。
- Discovery of Novel Gain-of-Function Mutations Guided by Structure-Based Deep Learning
- Authors: R. Shroff, A.W. Cole, D.J. Diaz, B.R. Morrow, I. Donnell, A. Annapareddy, J. Gollihar, A.D. Ellington, R. Thyer
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00345
- Institution: The University of Texas at Austin, USA
周辺情報からの残基の予測でそんなに上手く行くとは、モデルの訓練で機能に関しての情報は与えていないけど。
Metabolic/Signal Pathway Engineering
シグナル経路、代謝経路、酵素工学など- Adaptive laboratory evolution of native methanol assimilation in Saccharomyces cerevisiae
- Authors: M.I. Espinosa, R.A. Gonzalez-Garcia, K. Valgepea, M.R. Plan, C. Scott, I.S. Pretorius, E. Marcellin, I.T. Paulsen & T.C. Williams
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19390-9
- Institution: Macquarie University, Australia
- Awakening a latent carbon fixation cycle in Escherichia coli
- Authors: A. Satanowski, B. Dronsella, E. Noor, B. Vögeli, H. He, P. Wichmann, T.J. Erb, S.N. Lindner & A. Bar-Even
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19564-5
- Institution: Max Planck Institute, Germany
タイトルがかっこいい、内容もタイトルに劣らずかっこいい。
- Synthesis of Ligustrazine from Acetaldehyde by a Combined Biological−Chemical Approach
- Authors: K. Peng, D. Guo, Q. Lou, X. Lu, J. Cheng, J. Qiao, L. Lu, T. Cai, Y. Liu, H. Jiang
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00113
- Institution: Chinese Academy of Sciences, China
- Integrative Biosynthetic Gene Cluster Mining to Optimize a Metabolic Pathway to Efficiently Produce L‑Homophenylalanine in Escherichia coli
- Authors: Zhenning Liu, Dengwei Lei, Bin Qiao, Shilin Li, Jianjun Qiao, and Guang-Rong Zhao
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00363
- Institution: Tianjin University, China
代謝工学もin silicoで色々出てきて面白い。
- De Novo Biosynthesis of Multiple Pinocembrin Derivatives in Saccharomyces cerevisiae
- Authors: X. Liu, J. Cheng, X. Zhu, G. Zhang, S. Yang, X. Guo, H. Jiang & Y. Ma
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00289
- Institution: Chinese Academy of Science, China
こちらもin silicoでのRNA-seqデータのマイニング。
DNA / Biophysics
DNA、核酸論理回路、ナノスケール構造物など- Rapid and robust assembly and decoding of molecular tags with DNA-based nanopore signatures.
- Authors: K. Doroschak, K. Zhang, M. Queen, A. Mandyam, K. Strauss, L. Ceze & J. Nivala
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19151-8
- Institution: University of Washington, USA
バーコード配列を進化させるという発想が初耳で面白かった。
- DNA synthesis for true random number generation
- Authors: L.C. Meiser, J. Koch, P.L. Antkowiak, W.J. Stark, R. Heckel & R.N. Grass
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19757-y
- Institution: ETH Zurich, Switzerland
思ったよりバイアス大きくて驚き。
- Complex multicomponent patterns rendered on a 3D DNA-barrel pegboard
- Authors: S.F.J. Wickham, A. Auer, J. Min, N. Ponnuswamy, J.B. Woehrstein, F. Schueder, M.T. Strauss, J. Schnitzbauer, B. Nathwani, Z. Zhao, S.D. Perrault, J. Hahn, S. Lee, M.M. Bastings, S.W. Helmig, A.L. Kodal, P. Yin, R. Jungmann & W.M. Shih
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-18910-x
- Institution: Harvard University, USA
でかい、すごい。
Alternative Hosts / Strain Engineering
宿主や系統株の開発- Leveraging Oxidative Stress to Regulate Redox Balance-Based, In Vivo Growth Selections for Oxygenase Engineering
- Authors: S. Maxel, E. King, Y. Zhang, R. Luo & Han Li
- Journal: ACS Synthetic Biology
- Year: 2020
- DOI: 10.1021/acssynbio.0c00380
- Institution: UC Irvine, USA
directed evolutionのために株ごと作るという発想の転換。株そのものの汎用性は低いけど、アイデアは他に活かせそう。
Bioinformatics
- UMI-linked consensus sequencing enables phylogenetic analysis of directed evolution
- Authors: P.J. Zurek, P. Knyphausen, K. Neufeld, A. Pushpanath & F. Hollfelder
- Journal: Nature Communications
- Year: 2020
- DOI: 10.1038/s41467-020-19687-9
- Institution: University of Cambridge, UK
デミナーゼの進化についてデータベースも公開されているので、in silicoツール制作に使える。